原核生物と真核生物の転写調節の仕組みの違い
これまで「1)原核生物の転写調節の仕組み」と「3)真核生物の転写調節の仕組み」について学んできました。ここでは、真核生物と原核生物の転写調節の仕組みの違いについて、再確認していきましょう。
①プロモーター配列の違い
原核生物のプロモーター配列には、「-10領域(プリブノウボックス)(TATAAT)」と「-35領域(TTGACA)」と呼ばれる共通のコンセンサス配列(共通配列)が存在します。
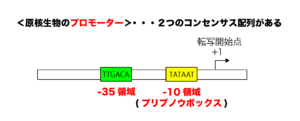
一方、真核生物のプロモーター配列には、「TATAボックス(TATAAAA)」と呼ばれるコンセンサス配列があります。
②RNAポリメラーゼの違い
原核生物のRNAポリメラーゼは1種類しかありません。
一方、真核生物のRNAポリメラーゼには3種類(PolⅠ、PolⅡ、PolⅢ)あります。
③プロモーター認識の違い
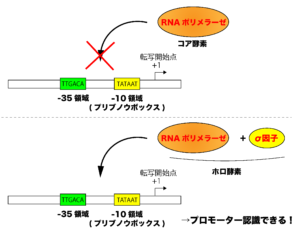
原核生物のRNAポリメラーゼは、プロモーター配列の認識に「σ因子(シグマ因子)」というタンパク質を必要とします。
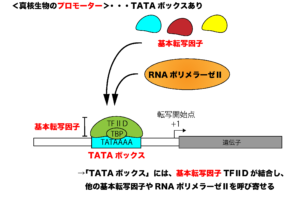
一方、真核生物のRNAポリメラーゼは、プロモーター配列の認識に複数の「基本転写因子」(TFⅡDなど)というタンパク質群を必要とします。
④転写調節因子の違い
大腸菌の「ラクトースオペロン」を想定していただくとわかりやすいかと思いますが、原核生物では転写調節因子(リプレッサーなど)がオペレーターに結合することによって転写調節をしています。
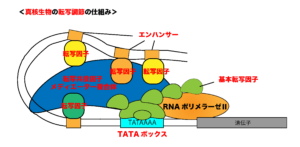
一方、真核生物では、複数の転写調節因子(単に「転写因子」と呼ぶことも多い)が、ある遺伝子のプロモーター配列に結合するとともに、その遺伝子のさらに上流(あるいは下流)にある「エンハンサー」や「サイレンサー」と呼ばれるDNA配列にも結合します。また、これらの転写制御配列がプロモーターから数千塩基以上離れて存在することもありますので、真核生物では転写調節の仕組みがより大規模であるといえます。
④転写共役因子の有無
真核生物には、転写因子や基本転写因子と共役することによって機能する転写共役因子(コファクター)と呼ばれるタンパク質群が複合体を形成することによって、転写調節をしていることも原核生物との大きな違いになります。
⑤クロマチン構造の有無
真核生物のDNAは、ヒストンと呼ばれる塩基性タンパク質に巻き付く形で「ヌクレオソーム」という構造体を形成しています。このヌクレオソームという構造体は真核生物に特有のもので、真核生物の染色体はこのようなヌクレオソームがいくつも数珠状につながった「クロマチン構造」をとっています。
このクロマチン構造は、ヒストンの化学修飾やDNAメチル化などの「エピジェネティクス」と呼ばれる制御の仕組みによって、弛緩と凝集が調節されていて、これが真核生物の転写調節における複雑な生体応答を可能にする仕組みの一つとなっています。
※エピジェネティクスを簡単にいうと「DNAの塩基配列の変化によらない遺伝子発現制御の仕組み(あるいはその研究分野)」のことをいいます。このような変化は、細胞分裂を経過しても引き継がれることが知られています。
⑥ポリシストロニック転写とモノシストロニック転写
原核生物の転写は、複数の遺伝子を1つのプロモーターの制御下でまとめて転写させるポリシストロニック転写と呼ばれる様式を採用しているという特徴があります。
原核生物と真核生物の転写制御の仕組みの違いについてはこれで以上です。
次は「1)RNAプロセシング(転写後調節)」について学んでいきましょう。
-
-
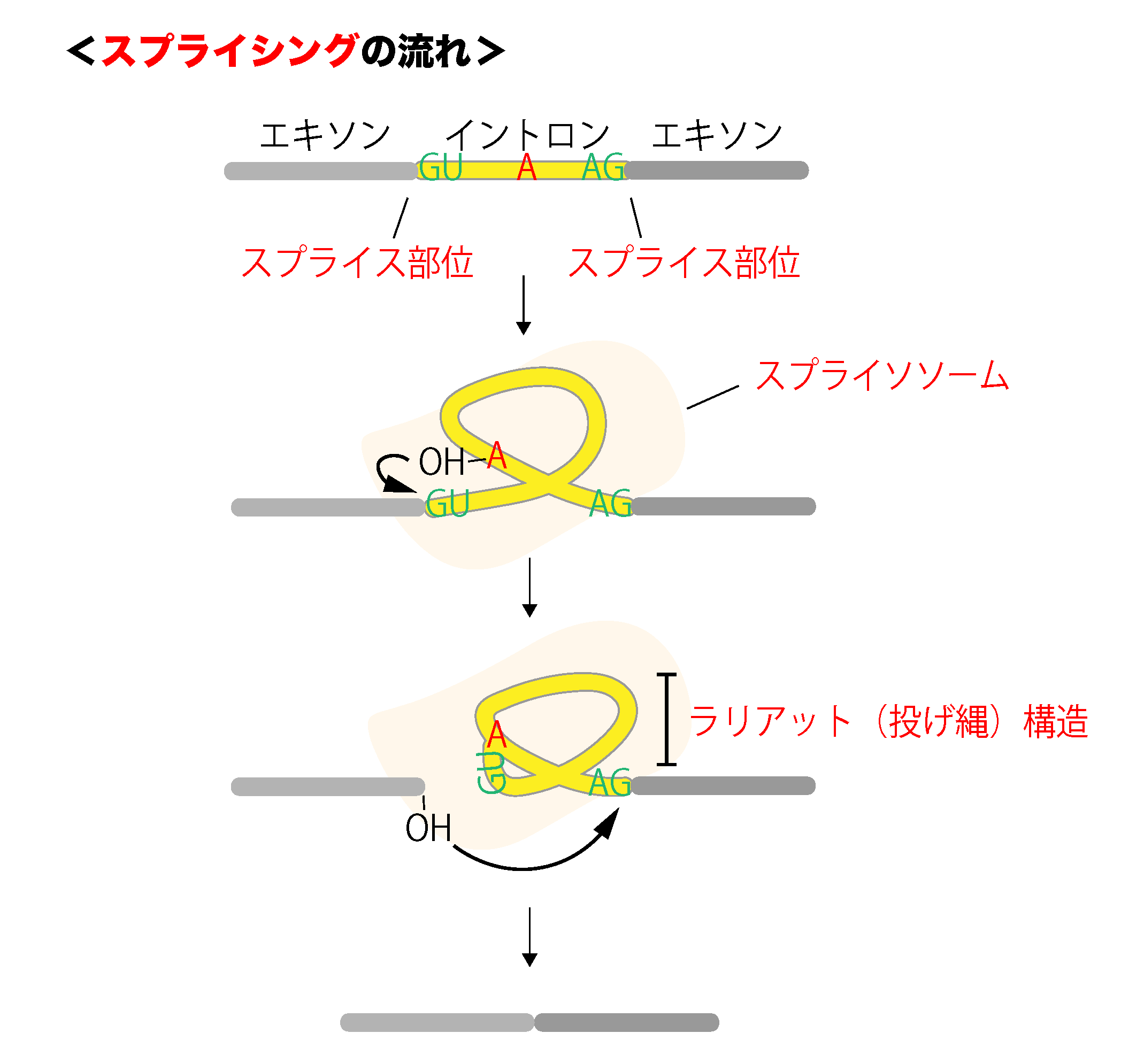
RNAのプロセシング(転写後調節)
RNAプロセシング 真核生物における遺伝子発現調節の仕組みの特徴の一つには、転写された前駆体mRNAが「RNAのプロセシング」という過程を経る、ということがあります。これらのプロセシングという過程は ...
続きを見る
【参考】


