真核生物の転写調節の仕組み
真核生物の遺伝子発現の調節には、さまざまな転写因子やその制御配列(エンハンサーやサイレンサー)が関与しています。ここでは、真核生物の転写調節の仕組みについて学んでいきましょう。
1.転写開始
真核生物のプロモーターには、「TATAボックス(TATAAAA)」と呼ばれるコンセンサス配列があります。真核生物では、このTATAボックスをもとに(TATAボックスが転写開始部位の目印となって)転写が開始されることが多いのですが、このTATAボックスを含むプロモーター領域には、RNAポリメラーゼが直接結合することはできません。
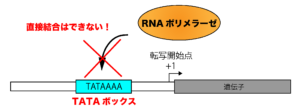
※TATAボックス=プロモーターではないので注意してください。
ここで覚えておきたい真核生物の特徴として、
ということが挙げられます。
基本転写因子の一つであるTFⅡDには、TBP(TATA結合タンパク質、TATA-binding protein)と呼ばれるサブユニットがあります。このTBPはその名の通り、TATAボックスに結合することができますので、転写開始時には、基本転写因子TFⅡDがTATAボックスに結合することによって、その他の基本転写因子やRNAポリメラーゼがプロモーター配列へと侵入してこれるようになり、mRNAの転写が開始されます。
真核生物のmRNAはRNAポリメラーゼⅡによって転写されますが、このとき、RNAポリメラーゼⅡのC末端領域(CTD:Carboxyl-Terminal Domain)がリン酸化されることによって、RNAポリメラーゼⅡの転写活性が高まります。
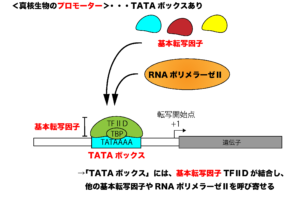
※TATAボックスは、原核生物のプロモーターにあるコンセンサス配列「-10領域(プリブノウボックス)(TATAAT)」に相当しますが、真核生物のTATAボックスは転写開始点から上流25-30bpにあるという違いがあります。
※原核生物の場合、RNAポリメラーゼが働くにはσ因子(シグマ因子)と呼ばれるタンパク質が必要でしたが、真核生物では代わりに複数の「基本転写因子」が必要であるというイメージを持たれても良いかもしれません。
2.真核生物のRNAポリメラーゼの種類
真核生物には、以下の3つのRNAポリメラーゼ(PolⅠ、PolⅡ、PolⅢ)がありますが、特に、mRNAの転写調節においては、RNAポリメラーゼⅡが関与しているということは覚えておきましょう。
・RNAポリメラーゼⅠ(PolⅠ)・・・rRNA合成
・RNAポリメラーゼⅡ(PolⅡ)・・・mRNA合成
・RNAポリメラーゼⅢ(PolⅢ)・・・tRNA合成、5S rRNA合成

3.転写調節(RNAポリメラーゼⅡによる転写)
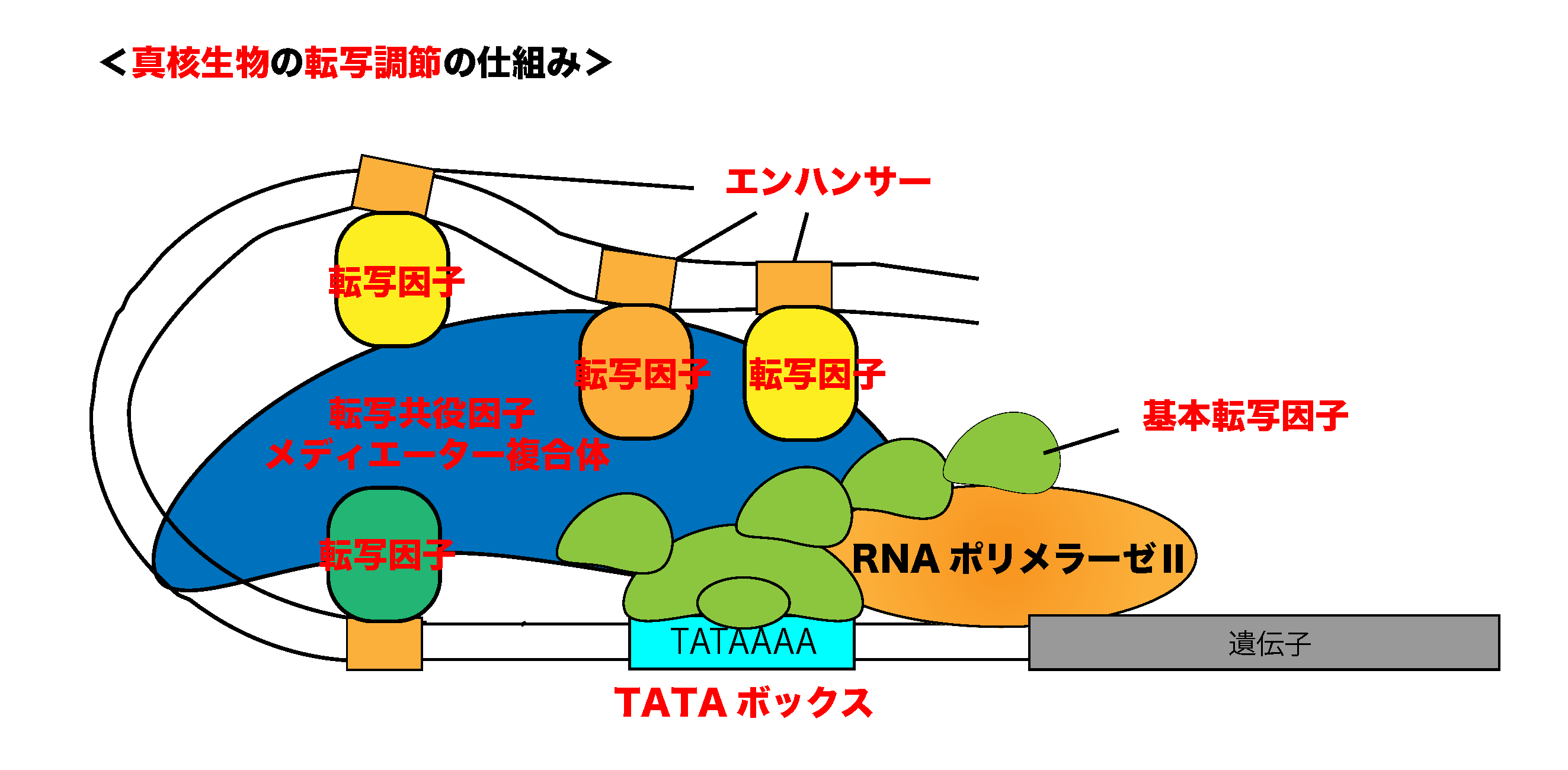
○複数の転写因子による調節
真核生物の遺伝子発現は、原核生物と同様に「転写調節因子(単に転写因子と呼ばれることが多い)」というタンパク質によって調節されています。この転写調節因子には、大きく分けて2つがあり、転写を正に制御するものを「アクチベーター」、転写を負に制御するものを「リプレッサー」といいます。
このような仕組みは原核生物にもあるのですが、
ここで覚えておきたいのは、真核生物では、複数の転写調節因子が一つの遺伝子発現の調節に関与しているということです。
※転写因子という用語は、DNAに直接結合することで転写を調節するタンパク質のことを意味しますので、正確には、転写因子というと「転写調節因子」と「基本転写因子」の両方を意味しています。
○エンハンサーとサイレンサー
上で述べたように、真核生物では複数の転写因子によって巧みに遺伝子発現の調節が行われています。これらの転写因子は、ある遺伝子のプロモーターに結合するだけではなく、遺伝子のさらに上流(あるいは下流)にある「エンハンサー」や「サイレンサー」と呼ばれるDNA配列にも結合します。
エンハンサーとは、転写を促進させる調節領域のことで、逆にサイレンサーとは、転写を抑制させる調節領域のことをいいます。
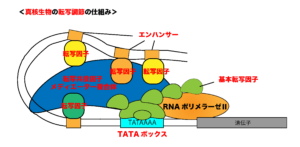
※これらの制御配列はプロモーターから数千塩基以上離れて存在することもありますが、プロモーターにも複数のエンハンサーやサイレンサー配列があり、そこに転写因子が直接結合します。
基本転写因子はすべての遺伝子の転写に共通して必須な制御因子なのですが、転写調節因子(いわゆる転写因子)にはさまざまな種類があり、それぞれ認識するDNA配列も異なっています。例えば、CREB(cAMP response element binding protein)という転写因子は、cAMP応答配列(CRE)というDNA配列に結合することによって標的遺伝子の発現を調節します。
また、さまざまな刺激に応じて、細胞内で機能する転写調節因子の組み合わせも違ってきます。そのため、これらの転写調節因子の組み合わせによって、どの遺伝子がどのタイミングで転写されるかは巧みに調節されています。
○転写共役因子(コファクター)による調節
「転写共役因子」とは、DNAには直接結合しないが、転写因子や基本転写因子とのタンパク質-タンパク質相互作用によって転写を調節するタンパク質のことです。転写因子や基本転写因子と共役することによって機能することから「コファクター」とも呼ばれます。
コファクターは、基本的には複合体を形成して働くことが知られているのですが、このとき転写の活性化に働くものを「コアクチベーター」といい、転写の不活性化に働くものを「コリプレッサー」といいます。
特に覚えておきたいのは、
ポイント
ヒストンアセチル化酵素(HAT、Histone Acetyl Transferase)やヒストン脱アセチル化酵素(HDAC、Histone Deacetylase)といったタンパク質が含まれている複合体では、それぞれ転写が促進、あるいは抑制される
ということです。
「ヒストンアセチル化とヌクレオソーム」で解説しているように、遺伝子発現というのは、クロマチン繊維の凝集と弛緩によって調節されています。簡単にいうと、ヒストンの化学修飾(アセチル化やメチル化)の変化によって、ヒストンとDNAの結合が弱くなったり、強くなったりします。
つまり、コアクチベーター複合体の中に、ヒストンアセチル化酵素(HAT)活性をもつタンパク質が存在すると、ヒストンがアセチル化されて、ヒストンの正電荷が消失することによって、ヒストンとDNAの結合が弱くなり、転写が促進されます。
逆に、コリプレッサー複合体の中に、ヒストン脱アセチル化酵素(HDAC)活性をもつタンパク質が存在すると、ヒストンが脱アセチル化されて、ヒストンとDNAの結合が強まり、転写が抑制されます。
また、メディエーター複合体と呼ばれるコアクチベーターの複合体も基本転写因子と転写調節因子をつなぐ重要なコファクターであることが知られています。
4.転写終結
真核生物のmRNAは転写された後には、3'末端に「ポリAテール」と呼ばれるアデニン(A)が20-200個も連続した配列が付加されるという特徴があります。
真核生物の転写の終結はあまりよく解明されていませんが、AAUAAA配列の下流20-30塩基においてmRNAの転写が終結し、その後すぐにポリAテールが付加されることによって、mRNAの安定化が行われます。
真核生物の転写調節の仕組みについてはこれで以上です。
次は「原核生物と真核生物の転写制御の仕組みの違い」について学んでいきましょう。
-
-
原核生物と真核生物の転写調節の仕組みの違い
原核生物と真核生物の転写調節の仕組みの違い これまで「1)原核生物の転写調節の仕組み」と「3)真核生物の転写調節の仕組み」について学んできました。ここでは、真核生物と原核生物の転写調節の仕組みの違い ...
続きを見る
【参考】

