原核生物の転写調節の仕組み
原核生物と真核生物では、遺伝子発現調節の仕組みが大きく異なっています。原核生物は真核生物ほど複雑な転写調節の機構をもちませんが、原核生物に特有の転写調節の仕組みというものがあります。ここでは、原核生物の転写調節の仕組みについて学んでいきましょう。
※遺伝子が発現するというのは、DNAにコードされているある遺伝子の情報がmRNAへと転写されて、機能をもつタンパク質へと翻訳されることをいいます。原核生物の場合は、この転写と翻訳が同時に進行することが特徴の一つになっています。
1.転写開始
原核生物のプロモーターには、「-10領域(プリブノウボックス)(TATAAT)」と「-35領域(TTGACA)」と呼ばれる共通のコンセンサス配列(共通配列)が存在します。
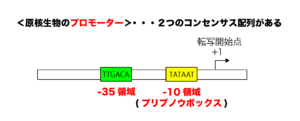
このプロモーター配列には、RNAポリメラーゼ(RNA合成酵素)と呼ばれる酵素が結合することによって転写が開始されていきますが、原核生物のもつRNAポリメラーゼ単体(コア酵素)には転写活性はあるのですが、プロモーター配列を認識する能力はありません。
原核生物では、このRNAポリメラーゼのプロモーター配列の認識にσ因子(シグマ因子)というタンパク質が必要になります。σ因子はコア酵素(RNAポリメラーゼ)に結合することによってプロモーター配列を認識できるRNAポリメラーゼ(ホロ酵素)に変換し、RNAポリメラーゼがプロモーター配列を認識できるようにします。
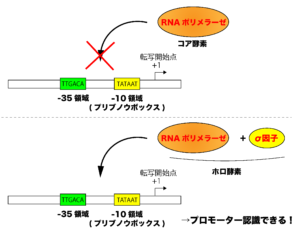
※σ因子は、一旦転写が開始されるとRNAポリメラーゼから解離して、他のσ因子をもたないコア酵素(RNAポリメラーゼ)に結合します。
2.転写調節
原核生物の転写は、ポリシストロニック転写と呼ばれる方法を採用しているという特徴があります。これは、複数の遺伝子を1つのプロモーターの制御下でまとめて転写させる様式のことです。
原核生物の遺伝子の上流には、転写の制御配列やプロモーター配列に加えて、オペレーターと呼ばれる配列があります。オペレーターには、転写の制御配列から作られたリプレッサーなどのDNA結合性因子が結合することによって、遺伝子の転写が巧みに調節されています。
これらのプロモーターやオペレーターなどの制御配列、さらにこれらによって制御される複数の遺伝子群を一単位で捉えたものを「オペロン」といいます。
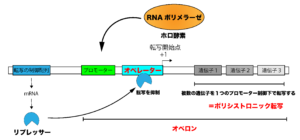
※特定の要因(熱などの刺激)によって誘導される調節因子(転写因子など)によって制御されるオペロンの一群のことを特に、レギュロンといいます。
3.転写終結
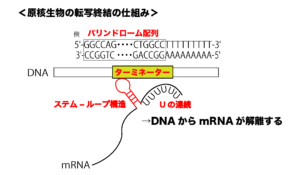
原核生物の遺伝子の下流には、転写を終結させるための「ターミネーター(転写終結配列)」と呼ばれる配列があります。
この転写終結のためのターミネーターの領域は、パリンドローム配列(回文配列、例:5'-GGCCAG・・・CTGGCC-3')と呼ばれる配列になっていることが多く、転写されたRNAはヘアピン構造やステム-ループ構造をとります。
※回文とは、「しんぶんし」のように、始めから読んだ場合と終わりから読んだ場合で同じ意味になる文のことです。
このようにRNAがヘアピン構造やステム-ループ構造をとると、物理的にDNAとの距離が離れて、転写されているRNAが解離しやすくなります。GとC(水素結合:3つ)が多く含まれるパリンドローム配列では、強固なヘアピン構造やステム-ループ構造が形成されます。
さらに、ターミネーターの領域の後ろから転写された部分には、ウラシル(U)が連続した領域が存在します。転写されたRNAにUの配列が連続すると、DNAとの結合が弱まり(AとUは水素結合:2つ)、転写されているRNAがより解離しやすくなります。
原核生物の転写調節の仕組みについてはこれで以上です。
次は「2)ラクトースオペロンと青白選択」について学んでいきましょう。
-
.png)
-
ラクトースオペロンと青白選択
ラクトースオペロンと青白選択 前項では「原核生物の転写調節の仕組み」について学びました。ここでは、原核生物の転写調節の例として、ラクトースオペロンの仕組みを学んでいきましょう。ラクトー ...
続きを見る