ラクトースオペロンと青白選択
前項では「原核生物の転写調節の仕組み」について学びました。ここでは、原核生物の転写調節の例として、ラクトースオペロンの仕組みを学んでいきましょう。ラクトースオペロンの原理は、実際に研究にも応用されていて、この手法は目的の遺伝子を導入した大腸菌を選択する「青白選択(ブルーホワイトセレクション)」という操作として広く知られています。
1.ラクトースオペロンとは
ラクトースオペロンとは、大腸菌などの細菌がもっている「ラクトースをエネルギー源として利用するための一連の酵素(ラクトース分解酵素など)の発現を制御しているオペロン」のことをいいます。オペロンについて「原核生物の転写調節の仕組み」で解説していますのでご確認ください。
※ラクトース(乳糖)はグルコース1分子とガラクトース1分子からなる二糖類のことです。
ラクトースオペロンには、LacZ、LacY、LacAの3つの遺伝子がありますが、このうちLacZはラクトースをグルコースとガラクトースに加水分解するβ-ガラクトシダーゼというタンパク質をコードしていることは覚えておいてください。
大腸菌は、普段はグルコースをエネルギー源として利用していますが、グルコースが欠乏した条件下では生存のために、ラクトースの利用が活発になります。
つまり、グルコースの存在下では、ラクトースを利用するためのラクトースオペロンは「off」の状態にありますが、グルコース欠乏かつラクトースの存在下では、ラクトースオペロンを「on」の状態に変化させる仕組みを大腸菌はもっているのです。以下では、この「ラクトースの存在下でのラクトースオペロンの調節」と「グルコース欠乏時のラクトースオペロンの調節」に分けて解説していきます。
○ラクトースの存在下でのラクトースオペロンの調節
ラクトースが存在しないときには、ラクトース合成酵素などを発現させる必要はありませんので、ラクトースオペロンは「off」の状態にあります。この時、転写の制御配列から作られたリプレッサーがオペレーターに結合することによって、転写が抑制されています。
-300x152.png)
-300x150.png)
○グルコース欠乏時のラクトースオペロンの調節
グルコースが存在するときには、ラクトースが存在していてもラクトースの利用は抑制されて、グルコースが利用されます。これは「グルコース効果」あるいは「カタボライト抑制」と呼ばれます。
グルコースは細胞内cAMP(サイクリックAMP)濃度を下げることによって、「CAP(cAMP activated protein)」というアクチベーターの活性化を抑制します。これによってRNAポリメラーゼのプロモーターへの結合が抑制されて、転写が抑制されます。
-300x158.png)
-300x158.png)
以上をまとめますと、
2.青白選択(ブルーホワイトセレクション)とは
青白選択(ブルーホワイトセレクション)とは、目的の遺伝子を導入した大腸菌を選択する手法のことです。例えば、ある遺伝子を制限酵素を使ってプラスミドベクターにライゲーション(挿入)するとき、すべてのプラスミドベクターに目的遺伝子を導入できる訳ではありません。中にはセルフライゲーションといって、目的遺伝子が導入されないまま、環状のプラスミドDNAに戻ってしまうこともあります。
そこで青白選択では、まず大腸菌のラクトースオペロンの「プロモーター」「オペレーター」「LacZ遺伝子(β-ガラクトシダーゼをコードしている)」の一単位をプラスミドベクターに組み込んだものを使用します。そして、このLacZ遺伝子の内部には、マルチクローニングサイト(MCS)という制限酵素によって認識される塩基配列が集まった部位が存在します。目的遺伝子をプラスミドベクターに導入するとき、このマルチクローニングサイトに目的遺伝子を導入します。
そのため、目的遺伝子を導入できたプラスミドベクターでは、LacZの遺伝子が2つに分断され、正常なβ-ガラクトシダーゼが発現されなくなります。
青色選択では、このような仕組みを利用して、まずIPTGと呼ばれるアロラクトース様の誘導物質を加えることによって、オペレーターに結合しているLacリプレッサーを阻害して、LacZの遺伝子発現を促進します。このとき、目的遺伝子を導入できなかったプラスミドベクターでは、LacZ(β-ガラクトシダーゼ)の遺伝子が正常に発現されます。そのため、β-ガラクトシダーゼの基質となり、分解されることで青色を呈するX-galという試薬を加えることによって、コロニーの色は青色に変化します。
一方で、目的遺伝子を導入できたプラスミドベクターでは、LacZの遺伝子が分断され、正常なβ-ガラクトシダーゼが発現しなくなることから、X-galは加水分解されず、白色のままになります。
-300x176.png)
したがって、IPTGとX-galを加えることによって、形成されたコロニーの色が白色であれば目的遺伝子が挿入されているプラスミドをもつ大腸菌であるということがわかりますので、このコロニー由来の大腸菌をLB液体培地で増殖させればよいということになります。
※IPTGはアロラクトースとは違い、β-ガラクトシダーゼ(LacZにコードされるタンパク質)の基質とはなりませんので、IPTGを加えるとβ-ガラクトシダーゼの転写が持続的に活性化できます。
ラクトースオペロンと青白選択についてはこれで以上です。
次は「3)真核生物の転写調節の仕組み」について学んでいきましょう。
-
-
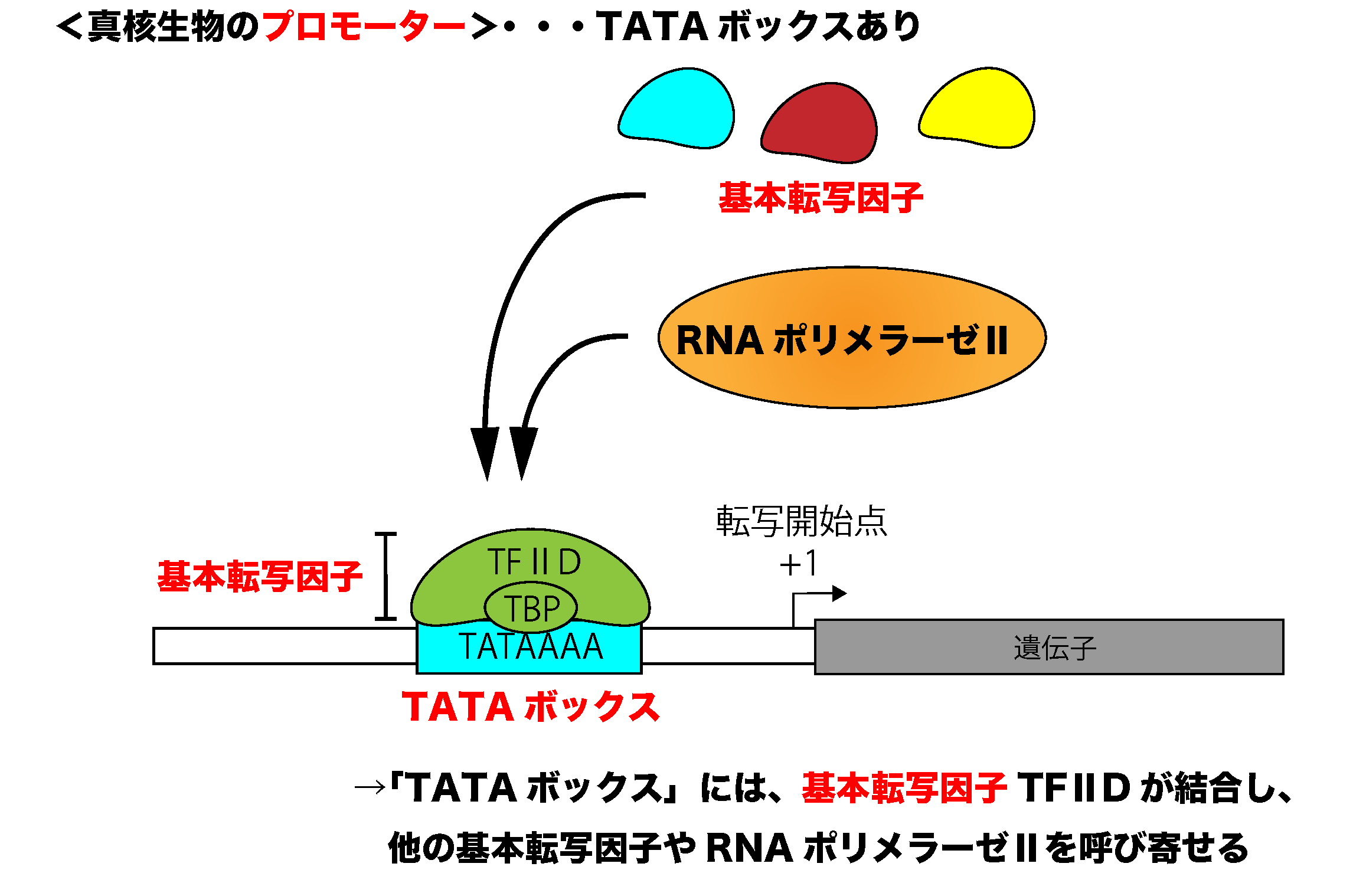
3)真核生物の転写調節の仕組み
真核生物の転写調節の仕組み 真核生物の遺伝子発現の調節には、さまざまな転写因子やその制御配列(エンハンサーやサイレンサー)が関与しています。ここでは、真核生物の転写調節の仕組みについて学んでいきまし ...
続きを見る